Rice Ragged Stunt Virus (Rrsv) – Virus Kerdil Hampa Padi

Sejarah
Virus Kerdil Hampa Padi (RRSV) pertama kali didokumentasikan pada tahun 1976-1977 di Indonesia dan Filipina [1,2]. Tak lama setelah itu, virus ini meluas menjadi wabah di banyak negara di Asia Selatan dan Asia Tenggara [3]. RRSV juga teridentifikasi di Taiwan pada tahun 1977 [4], di Tiongkok pada tahun 1978 [5], dan di Jepang pada tahun 1979 [6]. Asal-usul virus ini di wilayah-wilayah tersebut masih tidak pasti, meninggalkan pertanyaan apakah RRSV telah lama ada tanpa disadari atau telah menyebar dari tempat lain sebelum ditemukan. Wereng cokelat, Nilaparvata lugens Stal., berfungsi sebagai pembawa virus ini secara persisten [1,2]. Serangga ini dikenal karena kecenderungan migrasinya yang jauh, bahkan mereka dapat menyeberangi lautan [7]. Di daerah di mana wereng tersebut tidak dapat bertahan di musim dingin, serangga migran ini muncul sebagai sumber utama RRSV.
Sejak awal tahun 1970-an, populasi wereng cokelat mengalami peningkatan di berbagai negara di Asia Selatan dan Asia Tenggara. Peningkatan jumlah wereng ini disertai dengan eskalasi kerusakan yang disebabkan baik oleh wereng itu sendiri maupun oleh virus yang dibawanya, seperti Virus Kerdil Hampa Padi (RRSV) dan virus lain yang dikenal sebagai Virus Kerdil Rumput Padi (RGSV). Di Indonesia, kerugian yang diestimasi pada tanaman padi akibat wereng cokelat dan virus yang ditularkannya mencapai tingkat yang signifikan. Secara khusus, pada tahun 1975, 1976, dan 1977, kerugian mencapai 0,7, 1,1, dan 1,1 juta ton metrik beras kering tanpa sekam, masing-masing. Angka-angka ini setara dengan sekitar 3%, 3%, dan 5% dari total produksi padi di Indonesia selama tahun-tahun tersebut [8].
Kejadian RRSV secara utama dipengaruhi oleh kepadatan vektor utamanya, wereng cokelat, dan keberadaan sumber penyakit. Di daerah-daerah tropis Asia di mana budidaya padi terjadi sepanjang tahun, baik kejadian RRSV maupun populasi wereng cokelat cenderung tinggi. Daerah tropis menyaksikan migrasi terus-menerus wereng cokelat, memfasilitasi transfer RRSV dari daerah fokus ke daerah pertanian padi lainnya. Meskipun wereng cokelat dapat dikelola melalui aplikasi insektisida yang tepat, mengendalikan RRSV dan penyakit kerdil rumput menjadi tantangan, terutama di daerah dengan kepadatan wereng tinggi. Penggunaan insektisida yang tidak memadai bahkan dapat menyebabkan kembalinya wereng cokelat. Untuk mengatasi hal ini, disarankan untuk menanam varietas yang tahan terhadap wereng dan virus yang ditularkannya di daerah di mana kepadatan wereng umumnya tinggi. Selain itu, telah diusulkan strategi yang melibatkan penanaman secara bersamaan dan eradikasi tanaman padi selama beberapa bulan.
Morfologi
Partikel RRSV, saat diperiksa dalam sel tanaman dan vektor, menunjukkan bentuk bola dengan diameter sekitar 65 nm. Partikel ini mengandung inti yang berkepadatan elektron sekitar 45 nm dalam diameter, dikelilingi oleh selaput yang kurang padat dengan lebar sekitar 10 nm [3, 9]. Dalam beberapa kasus, partikel tersebut tampak polihedral dalam potongan, dan jarak antara pusat dalam susunan kristalin sekitar 68 nm. Preparat yang diwarnai negatif mengungkapkan partikel RRSV yang sebagian terdegradasi dan lebih kecil dari yang diamati dalam sel yang dipotong. Pewarnaan fosfotungstat menyoroti partikel bulat dengan diameter 55-60 nm, sedangkan pewarnaan asetat uranil menggambarkan partikel bulat dengan proyeksi atau duri [3,9,10,11,12,13].
Diameter tubuh partikel tersebut kira-kira 50 nm, dan duri membentuk bentuk kerucut terpotong, dengan ukuran sekitar 12 nm di bagian atas, 15-18 nm di bagian dasar, dan panjang 7-8 nm. Struktur filamen dengan konfigurasi heliks sering kali memanjang dari partikel-partikel tersebut [3,9]. Ketika partikel difiksasi dengan glutaraldehida atau paraformaldehida sebelum pewarnaan fosfotungstat, duri dan kadang-kadang ekstrusi struktur filamen juga diamati. Partikel-partikel berduri ini menunjukkan ukuran dan bentuk yang mirip dengan partikel subvirus dari Fijivirus.
Namun, keberadaan pasti RRSV dengan selubung luar yang lengkap, seperti yang terlihat pada Fijiviruses, masih tidak pasti. Potongan tipis partikel RRSV seringkali tampak polihedral dengan selubung luar, dan dalam pewarnaan asetat uranil, partikel kadang-kadang dikelilingi, sebagian menyerupai Fijiviruses [3,9]. Di sisi lain, Milne mengusulkan bahwa duri yang lebar pada beberapa profil mungkin memberikan tampilan selubung yang kontinu [12]. Sampai saat ini, virion ganda selubung lengkap belum diamati dalam persiapan RRSV dengan pewarnaan negatif [3,9,10,11,12].
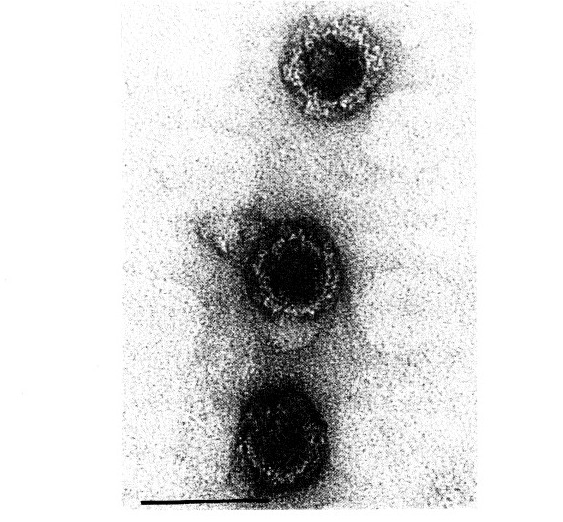
Gambar 1 Partikel RRSV dalam preparat gall tissue dari tumbuhan padi yang terinfeksi. Garis horizontal menunjukkan ukuran 0.1 mikrometer
Gejala dan Cakupan Inang
Bibit padi yang terinfeksi menunjukkan gejala infeksi Virus Kerdil Hampa Padi (RRSV) kira-kira 2-3 minggu setelah inokulasi [1,2,3]. Pertumbuhan daun baru terhambat, dan daun-daun ini cenderung lebih pendek, lebih sempit, dan berwarna hijau gelap dibandingkan dengan daun yang sehat. Seringkali, daun-daun tersebut menunjukkan deformitas, tampak hancur atau berbelit-belit. Seiring tanaman bertumbuh, jumlah daun yang hancur atau berbelit-belit umumnya menurun, dan gejala ini mungkin menjadi kurang terlihat pada daun-daun baru dua bulan setelah inokulasi.
Pada beberapa varietas, tanaman yang terinfeksi mungkin terlihat hampir sehat, kecuali untuk kerdil yang ringan dan daun yang kadang-kadang berbelit. Selama tahap awal pertumbuhan, jumlah anakan pada tanaman yang terinfeksi meningkat tetapi tetap lebih rendah daripada tanaman yang sehat pada tahap anakan berikutnya. Benjolan atau pembengkakan pembuluh terlihat di sepanjang urat daun pada permukaan luar selubung dan helai daun selama tahap anakan berikutnya. Gejala menjadi lebih jelas lagi pada tahap berdaun.

Gambar 2 Kiri: daun yang pendek, hancur, atau berbelit-belit pada tanaman yang terinfeksi Virus Kerdil Hampa Padi (RRSV). Satu bulan setelah inokulasi. Tengah: pembengkakan pada urat daun pada selubung tanaman yang terinfeksi RRSV. Kanan: penonjolan malai yang tidak lengkap dan daun bendera yang hancur dan berbelit-belit pada tanaman yang terinfeksi RRSV.
Daun bendera pada tanaman yang terinfeksi seringkali pendek, hancur, dan berbelit-belit. Penonjolan malai tertunda dan tidak lengkap, dan biji-bijian menunjukkan perubahan warna dan tetap tidak terisi. Anakan sering menghasilkan cabang nodal yang mungkin membawa malai kecil dan tetap berwarna hijau setelah berkepala. Di lapangan, tanaman padi sering terinfeksi baik oleh Virus Kerdil Hampa Padi (RRSV) maupun virus kerdil rumput. Tanaman yang terinfeksi ganda menunjukkan kombinasi gejala, termasuk daun yang hancur atau berbelit-belit yang menunjukkan gejala kerdil hampa, dan daun yang sempit, pendek, berwarna tidak normal dengan anakan berlebihan yang merupakan ciri khas kerdil rumput.
Diantara lima belas taksa padi yang diperiksa, gejala seperti kerdil, daun yang hancur, pembengkakan urat daun, dan cabang nodal diamati, menyerupai yang terlihat pada Oryza sativa [13]. Selain itu, saat jagung, barley, gandum, gandum hitam, oats, dan millet diinokulasi dengan RRSV, mereka menunjukkan gejala serupa dengan yang diamati pada padi [3,14]. Beberapa gulma juga diidentifikasi sebagai tanaman inang potensial untuk RRSV, sesuai dengan data yang belum dipublikasikan dari G.Z. Salamat dan H. Hibino. Namun, meskipun RRSV memiliki jangkauan inang yang lebih luas, inang tambahan ini mungkin tidak memainkan peran signifikan dalam epidemi RRSV. Hal ini karena tanaman-tanaman ini bukan inang dari wereng cokelat, kecuali untuk beberapa taksa padi tertentu. Selain itu, upaya untuk mendeteksi RRSV dari tanaman-tanaman ini melalui pemberian makan wereng cokelat cukup sulit atau tidak berhasil.
Gejala yang diinduksi oleh Virus Kerdil Hampa Padi (RRSV) bervariasi tergantung pada varietas tanaman. Sebagai contoh, pada varietas Sitopas, tanaman yang terinfeksi menghasilkan daun bendera yang normal panjangnya, dan 60% malai muncul secara sempurna [12]. Varietas yang toleran yang terinfeksi RRSV pada tahap bibit menunjukkan gejala pada tahap awal, tetapi gejala tersebut akhirnya menghilang, kecuali untuk pembengkakan urat dan kerdil yang ringan sesekali. Varietas ini kemudian berkembang untuk menghasilkan malai dengan biji yang terisi.
Hubungan antara RRSV dengan Sel dan Jaringan Tumbuhan
Partikel RRSV ditemukan dalam jaringan floem pada daun padi yang terinfeksi [9,14,15]. Partikel ini tersebar dalam sitoplasma atau terbenam dalam inklusi mirip viroplasma, yang terdiri dari filamen dengan diameter 7 hingga 10 nm dalam sel parenkim. Selain itu, partikel virus dapat terletak di dalam tabung saring, dan sel nekrotik dalam jaringan floem seringkali mengandung partikel ini. Jaringan gembalaan, yang terbentuk di sisi abaksial dari bungkus vaskular, disebabkan oleh hiperplasia jaringan floem [3,9,2]. Mikroskopi cahaya mengungkap inklusi amorf dalam beberapa sel jaringan gembalaan. Di bawah mikroskopi elektron, sel-sel gembalaan terlihat mengandung partikel virus dalam inklusi mirip viroplasma dalam sitoplasma [9]. Kadang-kadang, partikel virus membentuk larik dengan susunan kristalin. Sel-sel gembalaan yang terdegenerasi tersebar dengan partikel virus dan filamen, mungkin berasal dari inklusi, terlihat jelas. Namun, sel parenkim yang mengelilingi elemen xilem dalam jaringan gembalaan tidak mengandung partikel.
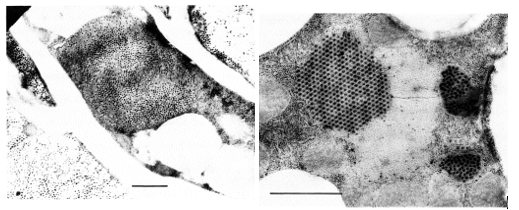
Gambar 3 Kiri: bagian sel gembalaan yang menunjukkan partikel RRSV tertanam dalam inklusi mirip viroplasma atau tersebar dalam sitoplasma yang terdegenerasi. Kanan: Bagian dari sel lemak yang menunjukkan agregat kristalin RRSV dan tabung serta vesikel fagositosis dengan partikel virus.
Upaya untuk mencegah infeksi Virus Kerdil Hampa Padi (RRSV) dengan mengendalikan Wereng Cokelat Padi (BPH) dengan insektisida dapat tidak sengaja memicu wabah RRSV. Dampak tidak disengaja ini seringkali disebabkan oleh dampak yang tidak proporsional dari insektisida terhadap musuh alami BPH. Insektisida tertentu, seperti triazophos, dapat meningkatkan tingkat kelahiran BPH dengan memengaruhi sistem reproduksi wereng jantan dan betina [16,17]. Lonjakan populasi BPH tidak hanya meningkatkan prevalensi RRSV dalam suatu lahan, tetapi juga mendorong transformasi BPH menjadi bentuk migrasi bersayap panjang (dikenal sebagai bentuk makropterus), memfasilitasi penyebaran RRSV ke lahan yang berjarak ratusan kilometer [18].
Selama wabah, membedakan antara kerugian hasil yang disebabkan oleh RRSV dan yang disebabkan oleh makanan BPH menjadi sulit. BPH juga berfungsi sebagai vektor untuk virus kerdil rumput padi (RGSV). Mengukur sejauh mana kerugian tanaman yang disebabkan oleh RRSV, RGSV, atau kerusakan akibat makanan BPH seringkali tidak praktis [19]. Uji lapangan yang dilakukan di Indonesia menunjukkan bahwa dengan kejadian penyakit kerdil hampa padi sebesar 34–76%, hasil gabah hanya mencapai 17–47% dari hasil tanaman yang sehat [20]. RRSV telah dilaporkan di beberapa negara, termasuk Cina, India, Indonesia, Jepang, Malaysia, Filipina, Sri Lanka, Taiwan, Thailand, dan Vietnam [21,22]. Keberadaan RRSV diduga, meskipun belum dikonfirmasi, di Kamboja, Laos, dan Myanmar. Secara mencolok, wabah besar RRSV dan RGSV terjadi di Cina dan Vietnam pada tahun 2006–2007 [23,24].
REFERENSI
[1] Hibino, R., Roechan, M. Sudarisman, S., and Tantera, D.M. (1977). A virus disease of rice (kerdil hampa) transmitted by brown planthopper, Nilaparvata lugens Stal. in Indonesia. Contr. Centr. Res. Inst. Agric. Bogor No. 35: 15 p.
[2] Ling, K.C., Tiongco, E.R., and Aguiero, V.M. (1978). Rice ragged stunt, a new virus disease. Plant Dis. Reptr. 62: 701-705.
[3] Hibino, H. (1979). Rice ragged stunt, a new virus disease occurring in tropical Asia. Rev. Plant Protect. Res. 12: 98-110
[4] Chen, C.C. and Chiu, R,J. (1981). Rice ragged stunt and its effect on the growth of rice plant. Plant Prot. Bull. (Taiwan, R.O.C.) 23: 67-75.
[5] Xie, L., and Lin, J. (1980). Rice ragged stunt disease, a new record of rice virus disease in China. Kexue Tongbao 25: 961-968.
[6] Shinkai, A., Nakano, M., and Iwasaki, M. (1980). Occurrence of rice ragged stunt (in Japanese). Ann. Phytopath. Soc. Japan 46: 411. (Abstr.).
[7] Kishimoto, R. (1971). Long distance migration of planthoppers, Sogatella furcifera and Nilaparvata lugens. Trop. Agric. Res. Ser. 5: 201-216.
[8] Mochida, 0. (1979). Brown planthoppers reduce rice production. Indonesian Agr. Res. Development J. 1(1-2): 1-7.
[9] Hibino, H., Saleth, N. and Roechan, M. (1979). Reovirus-like particles associated with rice ragged stunt diseased rice and insect vector cells. Ann. Phytopath. Soc. Japan 45: 228-239.
[10] 18. Kawano, S., Shikata, E., and Senboku, T. (1982). Purification and morphology of rice ragged stunt virus. J. Fae. Agre., Hokkaido Univ. 61 (Pt 2): 209-218.
[11] 19. Kawano, S., Uyeda, I., and Shikata, E. (1984). Particle structure and double stranded RNA of rice ragged stunt virus. J. Fae. Agr. Hokkaido Univ., 61 (Pt 4): 408-418.
[12] 25. Milne, R.G. (1980). Does rice ragged stunt virus lack the typical double shell of the Reoviridae? lntervirology 14: 331-336.
[13] IRRI (Int. Rice Res. Inst.). (1981). Annual Report 1980. p. 42-44.
[14] Chettanachit, D., Putta, M., and Disthaporn, S. (1978). Rice ragged stunt in Thailand. Int. Rice Res. Newsl. 3(4): 14-15.
[15] Shikata, E., Leelapanang, K., Tiangco, E.R., and Ling, K.C. (1977). Study indicates viral nature of rice ragged stunt disease. Int. Rice Res. News!. 2(5): 7.
[16] Ge, L.-Q., Yao Chen, J.-C. Wu, & Jahn, G. C. (2011). Proteomic analysis of insecticide triazophos-induced mating–responsive proteins of Nilaparvata lugens Stål (Hemiptera: Delphacidae). *Journal of Proteome Research*, DOI:10.1021/pr200414g.
[17] Wang, L. P., Shen, J., Ge, L.-Q., Wu, J.-C., Yang, G.-Q., & Jahn, G. C. (2010). Insecticide-induced increase in the protein content of male accessory glands and its effect on the fecundity of females in the brown planthopper, Nilaparvata lugens Stål (Hemiptera: Delphacidae. *Crop Protection, 29*, 1280–1285.
[18] Denno, R. F., Roderick, G. K., Olmstead, K. L., & Dobel, H. G. (1991). Density-related migration in planthoppers (Homoptera: Delphacidae): the role of habitat persistence. *The American Naturalist, 138*(6), 1513–1541.
[19] Hibino, R. (1979). Rev. Pl. Prot. Res. *12*, 98.
[20] Palmer, Soepriaman, & Mochida. (1978). Int. Rice Res. Newsl. *3*(3), 15.
[21] Antoniw, J. (n.d.). Show DPV and Refs in Frame. dpvweb.net. Retrieved December 17, 2016, from http://dpvweb.net
[22] Rice Knowledge Bank. (n.d.). Rice ragged stunt - IRRI. irri.org. Retrieved December 17, 2016, from http://irri.org
[23] Cabunagan, C., & Choi, H. (2009). Why outbreaks of virus diseases occurred in South Vietnam. Ricehoppers. Retrieved from http://ricehoppers.net/2009/06/why-outbreaks-of-virus-diseases-occurred-in-south-vietnam/
[24] Thomas, C. S., Nelson, N. P., Jahn, G. C., Niu, T., & Hartley, D. M. (2011). Use of media and public-domain Internet sources for detection and assessment of plant health threats. *Emerging Health Threats Journal, 4*, 7157. DOI:10.3402/ehtj.v4i0.7157
Artikel Lainnya
-
 Uncatgorized
UncatgorizedPanduan Praktis: Strategi Efektif untuk In-House Pest Management
-
 Uncatgorized
UncatgorizedJasa Profesional vs. Tim Internal: Mana yang Lebih Efektif untuk Pengendalian Hama?
-
 Uncatgorized
UncatgorizedKeuntungan Tim Internal untuk Pengendalian Hama dalam Bisnis
-
 Uncatgorized
UncatgorizedPengelolaan Hama Terpadu (PHT) Menjadi Solusi Terbaik dalam Menangani Hama di Sekitar Anda
-
 Uncatgorized
UncatgorizedMelindungi Standar Industri: Mengapa Pengendalian Hama Menjadi Kunci
-
 Uncatgorized
UncatgorizedPengendalian Hama Terpadu: Proses Berkelanjutan Jangka Panjang
-
 Uncatgorized
UncatgorizedProduk Aman, Bisnis Lancar: Dampak Hama terhadap Kualitas dan Reputasi di Industri Pengolahan Makanan
-
 Uncatgorized
UncatgorizedLindungi Reputasi & Keuntungan Bisnis Anda dari Serangan Hama!
-
 Uncatgorized
UncatgorizedHama Industri dan Hama Rumah Tangga: Perbedaan Karakteristik Yang Perlu Anda Ketahui
-
 Uncatgorized
UncatgorizedMakanan Favorit dalam Perilaku Makan Kecoa Jerman